In posts under “Our Shared Biology”, I will share small stories about the cool things cells do and how it connects to larger questions about our shared biology, questions that we might be compelled to discuss as scientifically literate citizens. I know I’m not the only one who thinks about these issues or has great ideas about them, so please reach out and share. I’d love to hear from you! (kbkaplan@ucdavis.edu)
Our shared cell biology: I study cells. When I meet someone who asks me what I do, I often struggle to find an answer that feels adequate. Saying “I study cells” tends to end conversations pretty fast. So, instead I say that I study the cellular basis for disease; you know, like how normal cells become cancer cells. This is true enough and the mention of cancer, which touches so many, can start important conversations. Still, I can’t help feeling afterward that I’ve sold myself — and cell biology more generally — a bit short. There’s a more honest and “preachy” answer to the question of “what do you do?” that percolates inside my head but never quite makes it way out. It might sound something like this: “I am a cell biologist. I study cells because knowing how they work inevitably leads me (and us) to the much larger story of who we are, where we came from and (maybe) where we are going. To tell this story, I try to understand how cells adapt to perturbations in order to maintain their identity, or fail to adapt in the case of disease. It is in understanding and ultimately connecting how cellular machines make cells so robust across the diversity of life on our planet that our shared biology can be slowly revealed, and we can begin to appreciate the enormous impact we humans have on this narrative. This is what I do.”
If that sermon didn’t make the point clearly enough, I can also offer up countless examples of our beautiful cell biology. My favorite this week? Did you know that blast-fungal pathogens — the bane of rice farmers across the world — use autophagy and septins to create a high pressure cellular “gun” that can penetrate the cell wall of a leaf? Crazy biology, right? The really crazy thing is that we use these same cellular machines in ways that might be mechanistically verrrrry similar. Cool, huh?
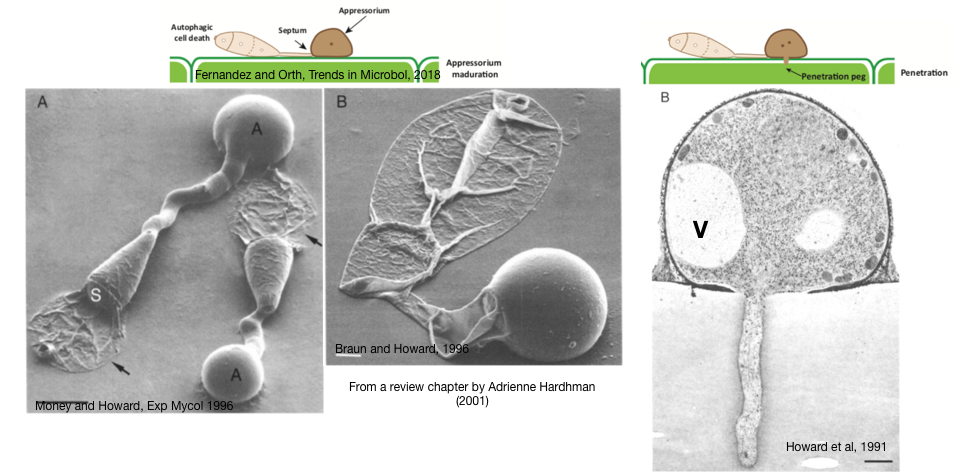
Blast Fungal Pathogens (for plants): The images below are scanning electron micrographs of one of these blast-fungal pathogens (Magnaporthe grisea, aka Magnaporthe Oryzea), This critter can grow as single cells, but under the right conditions comes together to make a three cell blast, or conidium. From the image below, you could be forgiven for thinking that a major part of the conidium has been “deflated”, because you’d be on the right track! The “stuff” inside the two “deflated” cells have in fact been degraded by a process called, autophagy (cell self-eating), where the contents of the cell are “shipped” off to the degradative organelle (i.e., vacuole or lysosome). In the middle image, you now see that the “germ tube” has also been deflated but the final area, the appressorium (A), does the opposite; it swells or inflates. In fact, the movement of all the degraded contents from the deflated cells of the conidium act to create pressure, which is funneled through to the appressorium and finally to a “penetration peg” (see the cartoons on top and the right hand image) that channels the turgor pressure (up to 8.0 MPa; 40X that of a car tire) to the growing peg in order to breakthrough the plant leaf and allow this alien-invader fungus to invade and spread INTO the plant cell. Penetration and invasion — yikes (think Alien, the movie)!
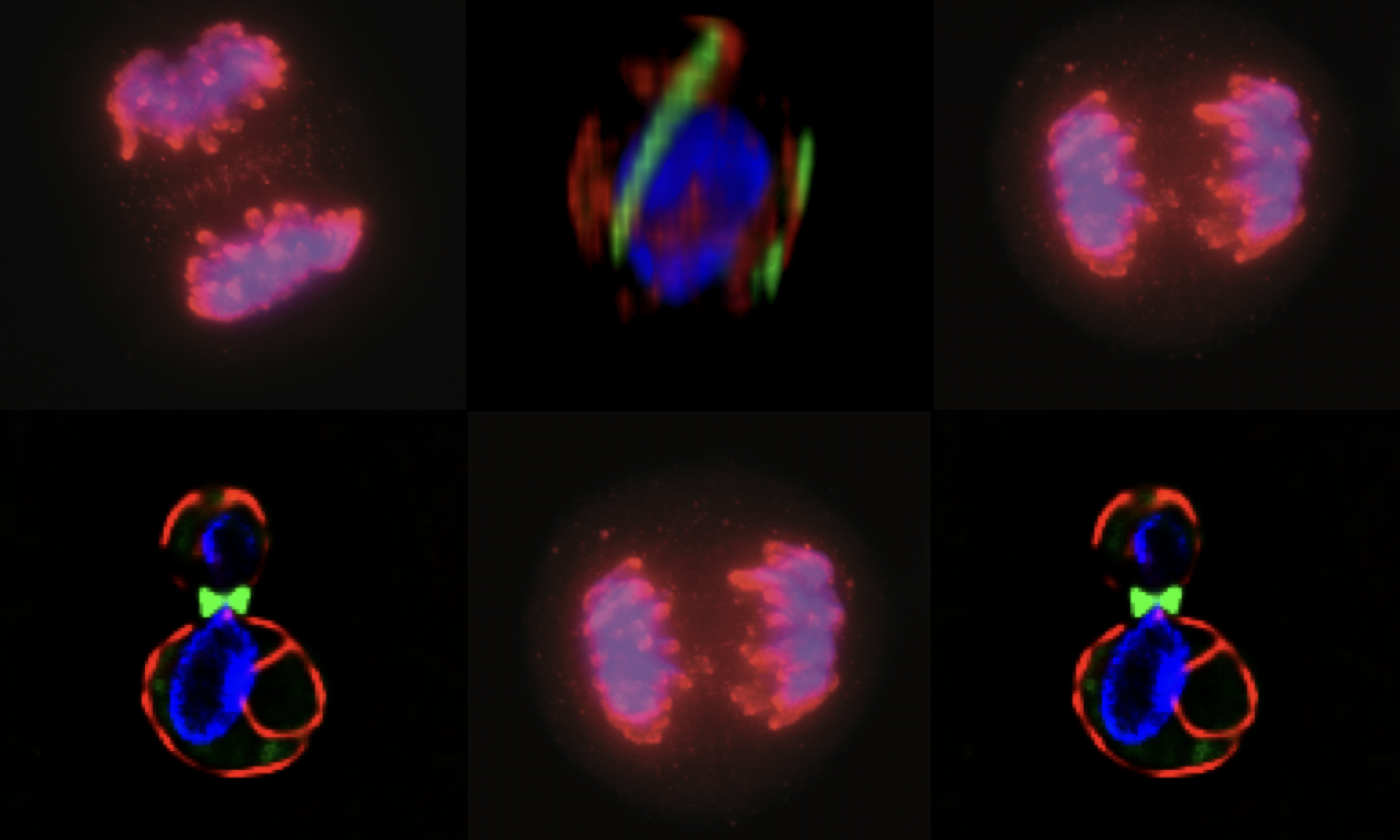
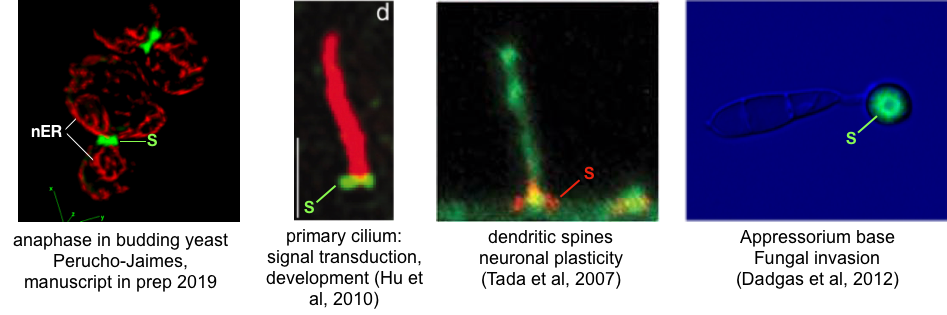
So what does this “Alien-like” biology have to do with our human-centric world view? Well, for one this blast-fungus is a major crop disease in rice and cereals, thus impacts the production of global food supply (not to mention having the potential to be a “weaponizable” biological weapon). We are also connected to this critter through the evolutionarily conserved cellular machinery that responds to environmental cues to organize the conidium into the invasion machine you see in the images. As the three cell conidium differentiates on the surface of the plant leaf, it begins to organize itself into distinct cellular regions by way of an understudied set of filaments called “septins”. Septins were first identified in the model organism budding yeast (S. Cerevisiae) where they were implicated in cell division. Since then, septins have been more broadly recognized to function in a diverse array of cell types in mammals. Septins are believed to be involved in forming physical barriers that regulate the diffusion of membrane proteins and contribute to the organization of such human-centric structures as primary cilia and dendritic spines, involved in key developmental signals and long-term potentiation in neurons, respectively (see example images below where “S” denotes septin structures). The rules that guide septin assembly in these circumstances remain unclear. Could a plant pathogen help us make sense of what is going on in our brains or during human development?
The answer to that question is “to be determined”, but the key idea here is that it SHOULD be determined. Fungal genetics has already provided some important clues as to the signals that are important for septin organization during appressorium development. The proteins that control appessorium development are recognizable to students of human biology as parts of canonical signaling modules (i.e., trimeric G protein-coupled receptors, Ras and MAP kinase cascades, and transcription factors) that control everything from the flight-fight response to immune function in normal and cancer cells. Figuring out how these cell signal modules regulate septins in fungal plant pathogens will undoubtedly tell us something about how these same modules work in our cells. Yet, making such a case to the National Institutes of Health to study biology in model organisms, much less in a plant pathogen, is more and more an excuse to not fund a grant as it doesn’t make it over the human relevance bar. I think this overly human-centric view on biology hurts our chances to truly understand the complexity of our own biology, a biology that is part of a 4 billion year story of life on Earth that has by random chance allows us to thrive. More generally, human-centric constructs have not generally served us or our planet well. If our society solely invests in biology that leads to the next pharmaceutical, we will be missing out on important biology that has truly transformative potential. I join a growing set of voices that advocate for bringing the modern tools of cell biology (molecular genetics, biochemistry and live cell imaging) to understanding the diverse manifestations of life on our planet as a window to truly see how our shared biology fits into the narrative of who we are and where we are going. What do you think? Email me and let me know.
For more on how blast-fungal pathogens work and how they are connected to our conserved cell biology, check out these cool articles:
Van Ngo, H. & Mostowy, S. Role of septins in microbial infection. Journal of Cell Science 132, jcs226266 (2019).
Hardham, A. R. in Biology of the Fungal Cell 10, 91–123 (Springer, Berlin, Heidelberg, 2001)